Der C-Kreislauf – ein neuer umfassender Ansatz!
Dr. Peter Vögele, Biologe
Zusammenfassung:
Das Standardmodell für den C-Kreislauf geht davon aus, dass die CO2-Konzentration der Atmosphäre seit 800 000 Jahren bis zu Beginn des Industriezeitalters niemals den Wert von 280 ppm überschritten habe und erst danach auf säkular einmalige Werte von ca. 400 ppm angestiegen sei (1). Ferner wird angenommen, dass der Austausch von CO2 zwischen Atmosphäre , Ozean und Biosphäre vor der Industrialisierung zu einem quasi stabilen Klimazustand geführt hätte, der erst durch die relativ kleine, aber zusätzliche menschliche Quelle aus der Kontrolle geraten sei (2). Der durch den Menschen verursachte Anteil an der globalen CO2-Emission wird auf lediglich 3% geschätzt, der aber über Hunderte von Jahren kumuliert, eine bedrohliche Prognose für die Zukunft liefern soll. Diese Prognose besteht erstens in einer globalen Temperaturerhöhung in Folge eines „Treibhauseffekts“ und zweitens in der „Versauerung“ des Meeres, weil ein Teil des zusätzlichen CO2 in Folge eines erhöhten Partialdrucks durch Diffusion auch in das Meer gelangt. Die dadurch verursachte Absenkung des pH-Wertes habe drastische negative Auswirkungen auf die Meeresorganismen (4,5).
Was ist falsch an diesem Modell?
- In der Erdgeschichte überwog der Verbrauch von CO2 durch die Photosynthese immer die CO2-Emission. Die Photosynthese hat eine CO2-Atmosphäre des Planeten in eine O2-Atmosphäre verwandelt. Ein „Gleichgewicht“ bzw. ein „stabiler Klimazustand“ sind fiktive Annahmen.
- Es ist pure Ideologie, den gegenwärtigen Anstieg des CO2 in der Atmosphäre auf einen marginalen anthropogenen Beitrag zurückzuführen. In einem guten Modell sollten alle, insbesondere die großen, Kohlenstoff-Flüsse berücksichtigt werden. Der schnelle gegenwärtige CO2-Anstieg (inclusive der C-Isotopenverschiebung) ist nicht einmalig.
- Das Modell ist statisch (Quellen, Senken), rein physikalisch und extrem simplifiziert. Die Ursachen der Kohlenstoff- Flüsse sind biologischer Art und variieren durch Rück-Kopplung, z.B. steigendes CO2 ergibt steigende Photosynthese. Die Änderungen der Flüsse gehen nicht in die Modelle ein.
- Der Austausch zwischen Meer und Atmosphäre wird von der biologischen und der physikalischen Pumpe getrieben. Die Annahme, dass der Austausch durch simple Diffusion wegen des marginal höheren Partialdrucks in Folge der anthropogenen Verbrennung stattfinde, ist nicht nur falsch, sondern lächerlich.
- Bei Abschätzung aller Flüsse ist es wahrscheinlich, dass der gegenwärtige CO2-Anstieg durch Emission aus dem Meer entsteht.
Erdgeschichtlicher CO2-Verlauf
Vor 3,8 Milliarden Jahren umfassten die Ozeane ca. 90% der Erdoberfläche, die wegen der ungehinderten (fehlende Ozonschicht) UVC-Strahlung ohne Leben war. Die Atmosphäre enthielt damals ca. 10% CO2 (100 000 ppm) und absolut keinen Sauerstoff. Heute enthält die Atmosphäre 21% Sauerstoff und nur noch 0,0% CO2. Diese Umwandlung der Erdatmosphäre ist das Werk eines einzigen Bakteriums ( Prochlorococcus), das die Blaulichtkomponente in Meerestiefen bis 200 m zur oxygenen Photosynthese ausnützt und so vor der UV-Strahlung geschützt ist. Dieses Cyanobacterium und sein Stickstoff-fixierender Artgenosse Synechococcus sind auch heute noch für 50% der globalen Photosynthese verantwortlich (6). Während ca. 1 Milliarde Jahren wurde der Sauerstoff, den Prochlorococcus produzierte, zur Oxidation von Eisen und Silizium verwendet, bis der Sauerstoffgehalt vor 2,2 Milliarden Jahren auch in der Atmosphäre anstieg. Die Ozonschicht entstand ca. vor 1,9 Milliarden Jahren (7).
Die weitere Entwicklung ab der Biologischen Explosion im Kambrium (vor 542 Millionen Jahren) zeigt die folgende Abb. (16)
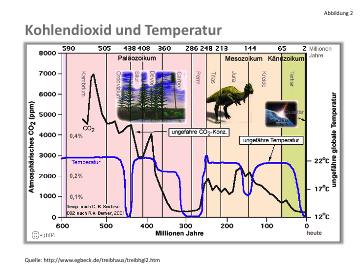
Vereinfacht gesehen, fällt die CO2-Konzentration bis nahezu Null am Ende des Carbon und steigt dann wieder auf 2500 ppm vor 150 Millionen Jahren an, um dann wiederum auf fast Null im heutigen Quartär zu fallen. Wenn man davon ausgeht, dass ab dem Kambrium zweifellos biologische Respiration (Atmung, Gärung, Zersetzung) stattfand, aber offensichtlich die Photosynthese (mit CO2-Verbrauch) überwiegt und deshalb ein „Gleichgewicht“ nicht konstruiert werden kann, dann stellt sich die Frage, wieso ein Boden der CO2-Konzentration bei ca. 150-200 ppm zweimal in der Erdgeschichte erreicht wurde. Das liegt daran, dass alle photosynthetisierenden Lebewesen bei niedrigem CO2-Gehalt der Umgebung Mechanismen zur Anreicherung entwickelt haben und ein Enzym Carboanhydrase, das die Umwandlung von gelöster Kohlensäure in gasförmiges CO2 katalysiert, weil Letzteres im Calvin-Cyclus gebraucht wird (8). Unter 200 ppm werden die Photosyntheseraten von C3 und C4-Pflanzen , von Algen und Bakterien extrem niedrig. Der starke Anstieg von CO2 im Perm wird auf vulkanischen Ausstoß zurückgeführt.
Bezüglich der Meeresversauerung in der Erdgeschichte folgen einige Abschätzungen von Zeebe (15) :
Vor 450 Millionen Jahren : 6000 ppm CO2 in der Luft und pH 7,4 im Meer
Vor 65 Millionen Jahren : 3000 ppm CO2 in der Luft und pH 7,6 im Meer
Vor 4,5 bis 2 Millionen Jahren: 525 – 200 ppm CO2 in der Luft und pH 8,0-8,3 im Meer.
Eisbohrkerne zur indirekten Ermittlung von CO2 (proxy-Methoden)
Vor etwa 2 Millionen Jahren begann das Eiszeit-Alter, in dem einer Eiszeit von ca. 100 000 Jahren eine Zwischeneiszeit von ca. 20000 Jahren folgt. Der Vorsitzende der Arbeitsgruppe 1 im IPPC , Prof. Stocker , ein Eiskernspezialist aus Bern, hat publiziert, dass am Ende einer Eiszeit erst die Temperatur steigt und mit einem Nachlauf von 800 Jahren erst das CO2 (9).
Während der Eiszeit liegen die CO2-Konzentrationen bei 200 ppm, in den Zwischeneiszeiten bei 280 ppm. Das impliziert, dass bei Beginn einer Zwischeneiszeit während mindestens 1000 Jahren ein CO2-Fluß aus dem Meer in Richtung Atmosphäre stattfand.
Um die Aussagekraft von Eisbohrkernen zu beurteilen, muss man wissen, dass die Umwandlung von Schnee über Firn bis zum Eis 200 Jahre dauern kann, in denen Diffusion von Gasen möglich ist. Eisbohrkerne können schnelle An-und Abstiege von CO2 innerhalb von 100- 500 Jahren nicht auflösen (10). Ferner ist der CO2-Gehalt in Polargebieten ca. 25 ppm niedriger wie Satellitenaufnahmen zeigen. Schnelle Änderungen des CO2-Gehalts können dagegen mittels der Analyse von Spaltöffnungen von Pflanzenmaterial entdeckt werden, z. B. ein ähnlicher Anstieg wie heute kurz vor der Jüngeren Dryas (von 150 auf 400 ppm in ca. 200 Jahren), der allerdings nicht zu einer Erwärmung, sondern zum Rückfall in eiszeitliche Temperaturen führte (11). Die Autorin führt den Anstieg auf „altes“ CO2 aus den Meerestiefen zurück, das eine ähnliche C-Isotopen-Verschiebung in der Atmosphäre bewirkte, wie auch heute zu beobachten ist.
Das IPCC –Modell der CO2-Austauschvorgänge ist falsch
In der Atmosphäre (750 Gt C) werden CO2- Emission durch Verbrennung, Respiration etc. und CO2-Aufnahme durch Photosynthese in wenigen Tagen durchmischt. Die Photosynthese übertrifft fast immer die Verbrennung und die Respiration , z. B. in den borealen Wäldern (26).
In den Ozeanen existiert eine gut ( von Wind und Wellen) durchmischte und warme Deckschicht von ca. 75 Metern (12), die von dem tiefen Wasser durch eine kältere Thermokline getrennt wird. Diese Deckschicht enthält 1000 Gt C und hat eine Durchschnittstemperatur im 20. Jahrhundert von 16,3 °C (im Gegensatz zur Landtemperatur von 11,1 °C). Die Temperatur der Ozeanoberflächenschicht hat sich in 150 Jahren um 0,7°C erwärmt nach Angaben der NOAA . Die Erwärmung der Oberflächenschicht des Meeres kommt vermutlich dadurch zu Stande, dass das Sonnenlicht bis zu 200 m tief eindringt und das Wasser erwärmt.
Der Austausch mit dem tiefen Ozean, der 50 mal mehr Kohlenstoff gespeichert hat, erfordert Zeiträume von 1000 Jahren.
Laut dem rein physikalischen Standardmodell von Sabine (17) erfolgt der CO2-Austausch zwischen Meer und Luft durch Diffusion, die vom leicht höheren Partialdruck in der Luft ( 2 ppm CO2 jährlich wegen der industriellen Verbrennung) getrieben wird. Die Austauschzeit wird auf 1 Jahr beziffert. Das Modell nimmt an, dass die höchste Konzentration von gelöstem anorganischem Kohlenstoff (DIC) wegen der Diffusion an der Meeresoberfläche vorliege. Das Modell berücksichtigt weder steigende Photosynthese in Folge steigender CO2-Konzentration noch den raschen Export von Kohlenstoff in den tiefen Ozean (18).
Im warmen Meer in Äquatornähe entweicht CO2 aus dem Meer , während es von kalten Meeren aufgenommen wird. In der folgenden Abb., die auf Daten des Scripps Institute beruht, wurde in jedem Meßpunkt die CO2-Konzentration in der Luft und im Wasser gemessen. Was überrascht, ist die Höhe der Differenz. In der Luft über den Warmwasserzonen wurden über 50 ppmv höhere CO2-Konzentrationen gemessen und über den kalten Zonen über 50 ppmv niedrigere. Das kann man nicht mit dem anthropogenen jährlichen Anstieg des atmosphärischen CO2 von 2ppmv erklären. Der CO2-Austausch zwischen Meer und Atmosphäre ist die Bilanz von 2 Flüssen: Emission aus dem Wasser in äquatorialen Zonen und Aufnahme aus der Luft in kalten Zonen (13). Eine globale Bilanz dieser beiden Flüsse kann nicht aus den Partialdrücken ermittelt werden (14), da bei diesem rein physikalischen Modell (17) der Effekt der biologischen Pumpe fehlt. In der Abb. sieht man, dass in dem kalten Teil des Meeres (links < 17°C) der CO2-Gehalt bei steigender Temperatur sinkt und nicht steigt, wie intuitiv zu erwarten wäre. Wegen der biologischen Pumpe (Absinken von Kohlenstoff in den tiefen Ozean) bildet sich ein vertikaler Gradient an anorganischem Kohlenstoff (DIC) aus, bei dem die C-Konzentration an der Meeresoberfläche am kleinsten ist. In kalten Meeren misst man mit dem Partialdruck im Wesentlichen die Stärke der biologischen Pumpe, in warmen Meeren die temperaturbedingte CO2-Ausgasung und in bestimmten „upwelling“ Zonen das Aufsteigen der ozeanischen Tiefenströmung. Allerdings liegen CO2-Konzentrationen über den „upwelling zones“ bei über 1000 ppmv.
Es bleibt offen, ob in der globalen Bilanz die Ozeane CO2 aufnehmen oder abgeben.
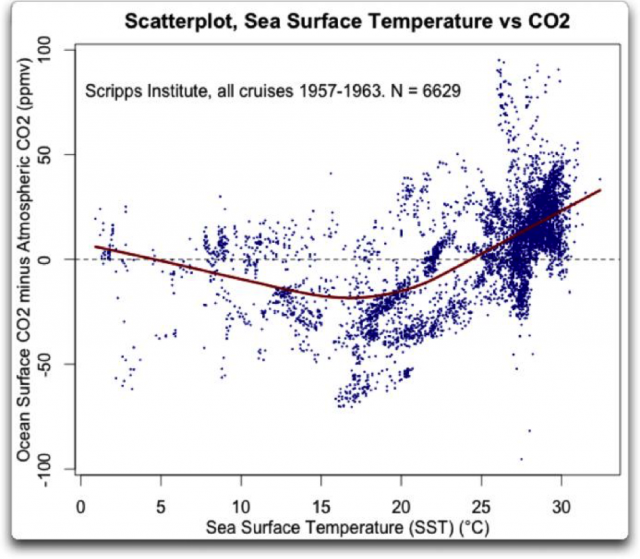
Quelle: NOAA, upwelling, pict.1
Photosynthese, wie funktioniert ein Korallenriff?
Die modellierte pH-Änderung im Great Barrier Reef seit 159 Jahren beträgt minus 0,054 pH (27). Im seichten Wasser eines Riffs schwankt der pH zwischen 8,4 am Tag und 7,7 bei Nacht (28). Die Abweichung des pH-Wertes nach oben während der Photosynthese ist einfach zu erklären. Das umgebende Wasser verarmt an anorganischem Kohlenstoff, der in Biomasse umgewandelt wird. Die tierische Respiration ist weit niedriger. Die Koralle besteht aus einem tierischen Organismus, z.B. einem Polypen, der eine geschlossene Kammer ausgebildet hat, in dem sich die photosynthetisierenden Algen befinden. Eine Protonenpumpe des Polypen senkt den pH der Kammer auf ca. 5,5, bei dem weitgehend nicht dissoziierte Kohlensäure vorliegt. Diese wird durch das Enzym Carboanhydrase (ähnlich wie im Blut) zu gasförmigem CO2 umgewandelt, das zur Bildung von Biomasse dient. Je stärker die Photosynthese, umso mehr verarmen die oberflächlichen Meereszonen an anorganischem Kohlenstoff und das Wasser wird alkalischer, nicht saurer. Auch die Kalkbildung ist kein chemischer Prozess, der von der Konzentration der Carbonat-Ionen abhängt, sondern ein biologischer Vorgang, der mit der Photosynthese korreliert ist, und unabhängig von pH-Schwankungen und Calcit-Sättigungswerten ist (30). Bei sinkendem pH-Wert sollte eigentlich die Carbonat-Konzentration und die Pufferkapazität abnehmen. In den Riffen tritt als negative Rückkopplung eine Kalkauflösung statt, die CO2 absorbiert und Carbonat freisetzt (29).
Gibt es ein Gleichgewicht der vorindustriellen CO2-Flüsse ?
Zunächst ein Zitat von Prof.Sabine (NOAA)(18) : „ although ocean biology plays an integral role in the natural distribution of carbon in the ocean, there is no conclusive evidence that the ocean uptake and storage of anthropogenic carbon, thus far, involve anything other than a chemical and physical response to rising atmospheric CO2.”
Der kurzfristige C-Kreislauf ist doch in erster Linie eine Bilanz zwischen biologischer Photosynthese und Respiration ( biologische Zersetzungsvorgänge mit Freisetzung von CO2) an Land und Meer, und in zweiter Linie der biologischen Pumpe. Welche physikalischen Vorgänge sollen denn eine Rolle spielen außer der marginalen anthropogenen Verbrennung?
Viele Organismen reagieren auf höhere CO2-Konzentration und höhere Temperatur mit einer erhöhten Photosyntheserate. Das Cyanobacterium Synechococcus (für 25% der globalen Photosynthese zuständig) erhöht die Photosyntheserate bei 750 ppm CO2 und 19°C auf das 4fache (19). Die quantitative Erhöhung der globalen Photosynthese (gross primary production) ist quantitativ schwierig zu fassen. Campbell (20) ermittelte 31% im 20.Jahrhundert. Im Sargasso Meer nahm das Mesoplankton um 61% zu zwischen 1994 und 2006 (21). Coccolithophoren nahmen zwischen 1990 und 2012 im Atlantik um 37% zu (22). Pro Jahr ergibt das Zunahmen von 0,31% bis 5,1 %.
Wenn die Brutto-Primärproduktion 223 Gt C pro Jahr beträgt (23), so müsste eine jährliche Abnahme von 0,7 Gt C bis 11,4 Gt C in der globalen C-Bilanz berücksichtigt werden. Dies liegt in der Größenordnung der anthropogenen Emission, wird aber in den Modellen nicht berücksichtigt.
Woodward (23) gibt folgende Zahlen in Gt C für Land und Ozean an:
GPP (gross primary production ) : Land 120, Ozean 103
NPP (net PP = GPP – pflanzliche Respiration): Land 60, Ozean 65
NET (net ecosystem prod. = NPP – heterotrophe (tierische) Respiration) : Land 5, Ozean 11
Man sieht, dass der photosynthetische C-Verbrauch immer die durch biologische Zersetzung emittierte C übertrifft, wie es auch in erdgeschichtlichen Zeiträumen der Fall war. Die Zahlen der NPP sind recht zuverlässig, da sie auf Satellitenmessungen der photosynthetischen Pigmente (400-700 nm) beruhen. Was nicht berücksichtigt wird, weil es von den Satelliten nicht erfasst wird, sind 25% der globalen Photosynthese durch Prochlorococcus in Meerestiefen bis 200 m. Die heterotrophe Respiration in den tropischen klaren Meeren kann in diesen Tiefen gleich Null gesetzt werden. Das bedeutet, dass ca 26 Gt C pro Jahr zusätzlich zur obigen Rechnung aus dem C-Kreislauf verschwinden. Das Cyanobacterium verdoppelt sich jeden Tag, aber die Hälfte der Zellen wird durch einen Phagen aufgelöst und endet im tiefen Ozean (biol. Pumpe).
Die NET im Ozean wird von Woodward mit 11 Gt C angegeben. Dieser Wert entspricht weitgehend der biologischen Pumpe, ist aber zu niedrig angesetzt. Laut Prof. Riebesell (24) liegt das Ausmaß der biologischen Pumpe bei 9-15 Gt C/ Jahr, liegt aber seiner Meinung nach eher beim oberen Wert. Man könnte folgende Bilanz des C-Kreislaufs aufstellen, wobei + einer Zunahme und – einer Abnahme von Kohlenstoff bedeutet:
Die NET (net ecosystem production) in Gt C/ Jahr) addiert sich aus -5 (Land) – 12-15 (Ozean) – 1 (jährliche Zunahme) , was 17- 21 Gt C / Jahr ergibt (ohne den Effekt von Prochlorococcus = 26) und die anthropogene Emission von 10 Gt C/ Jahr weit übertrifft. Das bedeutet, dass jedes Jahr der Atmosphäre trotz anthropogener Emission ca 7-11 Gt C entzogen werden. Wenn nur der halbe Effekt von Prochlorococcus (der von mir geschätzt wurde) mitgerechnet wird, ergibt sich ein CO2-Verbrauch von jährlich 30-34 Gt C trotz der „anthropogenen „ Emission.
Biologische Pumpe
Die biologische Pumpe ist so definiert, dass ein Teil (25%) des durch Photosynthese gebildeten organischen Kohlenstoffs relativ schnell und ohne heterotrophe Respiration in den tiefen Ozean absinkt, wodurch er dem C-Kreislauf für ca. 1000 Jahre entzogen wird (25). Am Meeresboden findet dann der Abbau des organischen Materials in anorganischen Kohlenstoff statt.
Das falsche Modell, das Prof. Sabine beim IPPC anwendet wird, beruht auf seiner falschen Einschätzung der Sinkgeschwindigkeit des organischen Materials : „ because anthropogenic CO2 invades the ocean by gas exchange across the air-sea interface, the highest concentrations of anthropogenic CO2 are found in near surface waters. Away from deep water formation regions, the time scales for mixing of near-surface waters downward into the deep ocean can be centuries“ (18).
Die Literatur gibt relative unterschiedliche Sinkgeschwindigkeiten an, aber man kann mit 30 bis 1000 Metern pro Tag rechnen, was mir von Prof. Riebesell (24) bestätigt wurde. Für Bakterien (Aggregate) werden Sinkgeschwindigkeiten von 440-660 Metren angegeben (18).
Wie oben diskutiert, wird der von Prochlorococcus produzierte Anteil der Nettoprimärproduktion von Satelliten nicht erfasst . Deswegen wird auch der gleich große Anteil an der Biologischen Pumpe einfach nicht berücksichtigt.
Die Biomasse landet am Boden der Ozeane , wo sie von Bakterien zu anorganischem Kohlenstoff abgebaut wird. Dieser Kohlenstoff kommt nicht vor 1000 Jahren wieder an die Oberfläche.
Physikalische Pumpe: Tiefenströmung , „upwelling“
Die physikalische Pumpe sorgt für den Austausch zwischen der Oberflächenschicht des Meeres und dem tiefen Ozean.
Die globale Meeresströmung (siehe Abb.) wird angetrieben durch die thermohaline Zirkulation. Der warme Golfstrom strömt an der Oberfläche des Atlantiks nach Norden und gibt Wasserdampf ab, der zu einer Abkühlung und Aufsalzung des Wassers führt. Auf der Höhe Islands ist die Dichte dieses Stroms so hoch geworden, dass er wie ein Wasserfall bis zum Meeresboden fällt. Dort verteilt sich die Strömung flächig und fließt sehr langsam nach Süden zurück. Die Zeit für einen Kreislauf wird auf ca. 1000 Jahre geschätzt.
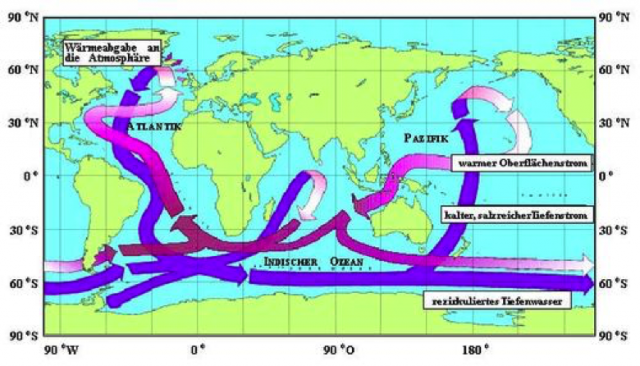
Das Austeigen von Tiefenwasser (upwelling) in Küstennähe wird dagegen hauptsächlich durch Winde verursacht, die die warme Oberflächenschicht wegtreiben und den Auftrieb des kalten und kohlenstoffreichen Tiefenwassers ermöglichen (30). In der Luft über den Auftriebsgebieten werden mehr als 1000 ppmv an CO2 gemessen (18).
Während der letzten Eiszeit war die Ventilation am Meeresboden unterbrochen, so dass sich dort Kohlenstoff ansammelte, der absolut frei von 14C war (31). Dieser Kohlenstoff kann bis zu 30 000 Jahren am Boden der Ozeane verbleiben (32).
Diskussion: woher kommt der gegenwärtige CO2-Anstieg in der Luft?
- Die durch Photosynthese und biologische Pumpe verursachte Aufnahme von CO2 aus der Atmosphäre bis in den tiefen Ozean, kann nicht durch anthropogene Verbrennung kompensiert werden. Die gemessene Zunahme in der Atmosphäre muss zwangsläufig durch Ausgasung aus dem Ozean erklärt werden.
- Entweder hat sich das „upwelling“ seit ca. 100 Jahren verstärkt, was am ehesten durch Änderungen der Windbewegungen erklärt werden kann oder die Tiefenströmung bringt seit 100 Jahren kohlenstoffreicheres Tiefenwasser nach oben.
- Der Befund, dass es sich um „alten Kohlenstoff“ (14C arm) handelt, könnte auch dadurch erklärt werden, dass CO2 aus dem Erdinneren austritt.
Quellen:
- IPPC Synthesis Report 2014, Summary for Polcymakers, SPM 1.2, Seite 4
- D. Schönwiese “Klimatologie”, 3.Aufl. Ulmer Verlag 2008, Seite 346
- Le Quére et al. 2014; Earth Syst. Sci. Data Discuss. 7, 521-610
- Schulz & U.Riebesell , S.160-162 in : J.Lozan (Hg): Warnsignal Klima: Die Meere-Änderungen & Risiken (2012)
- J. Hardt , C.Safina , Spektrum der Wissenschaft, Feb. 2011, 82-89
- Partensky et al. , Microbiol.Mol.Biol.Rev. 63, 106-127 (1999)
- Kutschera: Tatsache Evolution, München 2009,313
- Wang et al. , The Plant J. 82, 429-448 (2015)
- Monnin, T.Stocker et al, Science 291, 112-114 (2001)
- Van Hoof et al. , Tellus 57B, 351-355 (2005)
- Steinthorsdottir et al. , Quaternary Science Rev. 99, 84-96 (2014)
- Roedel “Die Atmosphäre”, 2.Aufl. 1994, Springer , Seite346
- Scripps Institute, Seawater CO2 Data ,published by C.D.Keeling 1957-1963
- Takahashi et al. 2016 , “Global Ocean Surface Water Partial Pressure of CO2 database. Oak Ridge Nat. Laboratory, U.S. Dep. of Energy
- E.Zeebe, Ann.Rev.Earth Planet Sci. 2012, 141-165
- A.Berner+ Z. Kothavala, Am.J.Sci 301, (2001), 182-204
- IPPC: Climate Change 2013, 6.3 (C.Sabine)
- Steele, https://wattsupwiththat.com/2017/03/02/how
- Fu, J.Phycology 43, (2007),485-496
- J.E.Campbell, Nature 544, (2017), 84-87
- Hernández-Léon et al., Limnol.Oceanogr. 55, (2010), 2503-2512
- K.M.Krumhardt et al. , Biogeosciences 13, (2017), 1163-1177
- I.Woodward, Current Biol. 17, (2007), 269-273
- Mitt. von Prof. U. Riebesell , Geomar, am 20.07.2017
- Körtzinger in : „Der marine Kohlenstoffkreislauf“ , S.16 ( Springer, 2017)
- Gonsamo et al., J.Geophys.Res. 122, 1275-1293 (2017)
- Wei et al. , J.Geophys.Res. 120, 7166-7188 (2015)
- I.Kline et al. , PLos ONE 10 (6): e0127648 (2015)
- W.Morse et al. , Chem.Rev. 107, 342-381 (2007)
- National Ocean Service, NOAA
- Basak et al. , Nature Geoscience 3, 770-773 (2010)
- L.Follett et al. , proc. Nat. Acad. Sci. U.S.A. 111, 16706 (2014)